AUTHOREA
Log in
Sign Up
Browse Preprints
LOG IN
SIGN UP
Ina Callebaut
Public Documents
1
June 05, 2020
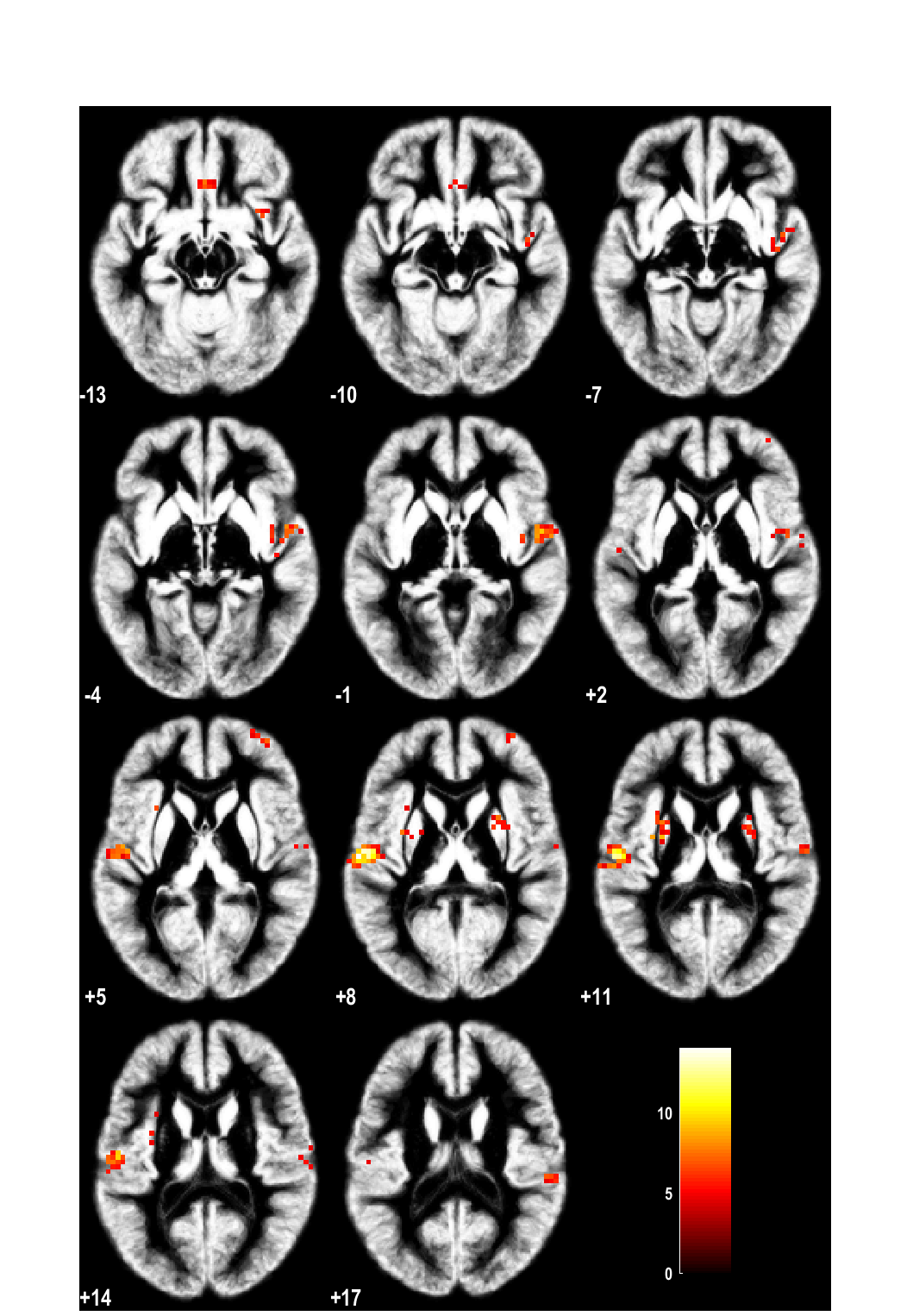
Brain activation after nasal histamine provocation in house dust mite allergic rhinit...
Ina Callebaut, Brecht Steelant, Wout Backaert, et al.