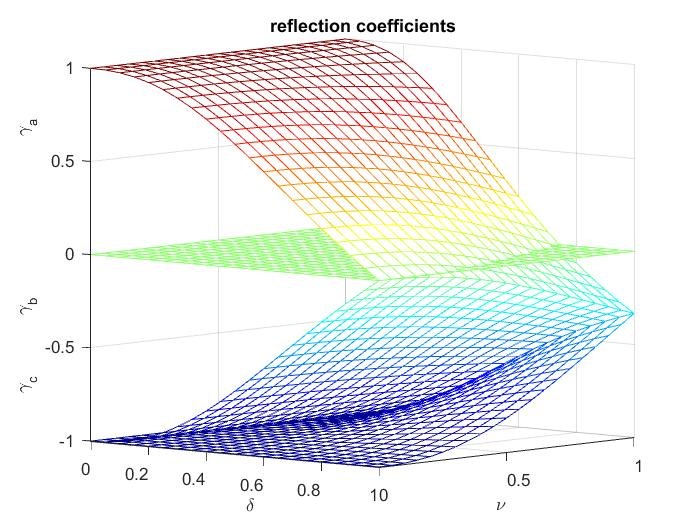
AbstractIntroduction[needs serious editing]The human systemic arterial system is too complex to describe in detail. Unlike the skeleton which has 206 bones in the normal adult, all of them named and located, the total number of arteries is unknown. There are probably one to two thousand named arteries (the exact number is very difficult to determine because surgeons and anatomists specialising in particular organs have named many arteries that are not named in general anatomical books. The number of unnamed arteries can only be estimated. One feature of the arterial tree is that it is primarily a bifurcating tree with relatively few anastomoses (loops in the teminology of network analysis). The well-known anastomoses are the Circle of Willis in the cranial circulation, the palmar and pedal arches in the hands and feet and a series of sequential anastomoses in the circulation of the intestines. There are certainly other anastomoses in the arterial tree buut they are the exception rather than the rule.One way to estimate the number of arteries is to estimate the number of capillaries in the body and then use the binary tree structure of the arteries to estimate the number of arteries necessary to service these capillaries. There are two separate ways of estimating the number of capillaries in the body from experimental measurements: an estimate based on the capillary density measured in various tissues and an estimate based on measurements of the volume flow rate through a capillary. We will show that both methods give similar values for the total number of capillaries in the body, given the variation of the different measurements. One stimate can be obtained by working back from the microcirculation where arterial blood is supplied via the arterioles (variously defined as 100-200 \(\mu\)m in diameter) through bifucating arteriol sphincters to the capillaries (5-10 \(\mu\)m in diameter) which form complex networks that depend on the tissue being perfused (parallel arcades in skeletal muscle, intricate networks with loops in the skin, etc.). Capillary density, technically the number of capillaries per unit volume of tissue but usually expressed as the number of capillaries per unit area of a thin section of tissue (or sometimes length of capillaries per unit volume), has been measured in a wide variety of tissues and these averages have been combined to estimate that there approximately 10 billion (\(10^{10}\)) capillaries in the human body. This estimate plus the assumption that the average length of a capillary is approximately 1 mm gives the oft-quoted statement that there are 25,000 miles of capillaries in the average body. If we define the smallest artery as the 'pre-arteriole' artery and assume that each pre-arteriole artery supplies 100 capillaries, then there must be \(10^8\) pre-arteriole arteries which means that there are \(2\times10^8\) arteries (in a uniform bifurcating tree there are \(2^N\) vessels in the \(N^{th}\) generation and \(2^N-1\) vessels in the generations leading up to the \(\)\(N^{th}\) gneration). This is almost certainly an over-estimate probably arising from the tendency to over-estimate when generating 'gee-whiz fact' for popular consumption..Another estimate of the number of arterries in the body can be obtained by estimating the number of generations of bifurcations there are in the arterial tree. A uniform binary tree with 20 generations would have \(2^{20}\sim2\times10^6\) arteries. This is probably an overestimate since, as discussed more fully below, the large systemic arteries (unlike the pulmonary arteries) do not form a full binar tree.Whatever estimate of the total number of arteries that we prefer, it is clear that larger named arteries represent only a tiny fraction of the total number of arteries. Since we cannot visualise, or even comprehend, the full arterial network it is essential to formulate a model that can be used to explore the properties of the the arterial system. This is the goal of this work. ------The cardiovasculoar system is composed of the heart and a complex network of vessels; large arteries, small arteries, the microcirculatory vessels (arterioles, capillaries and venules), the small veins and the large veins. The heart is divided into two parts, the left which pumps blood into the systemic arteries and receives blood from the pulmonary veins, and the right which pumps blood into the pulmonary arteries and receives blood from the systemic veins producing two separate circulatory systems, the systemic and the pulmonary. We will be concerned primarily with the systemic circulation and, unless stated, 'arteries' in the following will mean 'systemic arteries' but many of the ideas that will be discussed can be directly applied to the slightly simpler pulmonary circulation.The systemic circulation is important physiologically, perfusing all parts of the body except the lungs, and midically, with cardiovascular disease being the single largest cause of mortality world-wide. Arterial hemodynamics is a old field of research but despite extensive work and the importance of the subject much is still unknown.One problem is the anatomical complexity of the arterial circulation. In general the arteries have a tree structure, predominantly a binary tree, with relatively few loops. There are many examples of anastomoses (loops) in the circulation with the circle of Willis in the cranial circulation, the palmar arches in the hands and the plantar arches in the the feet being the best known. Another example of arterial anastemoses are the arteries lying on and perfusing the small ad large intestines. There are also a number of trifurcations in the circulation, notably the division of the lower aorta into the two iliac and coccugeal artery, particularly in animals with a tail. These loops and trifurcations may be important locally but they are relatively rare and we model the arteries as a binary tree without loops.The main problem with even this simple model of the arteries is the sheer number of arteries. The number of generations in the systemic arterial tree is estimated to be between 15 and 25 and a full binary tree with 20 generations will have \(2^{20}\ \sim10^6\) arteries. The number of named arteries is relatively small, probably a few thousand, and so most of the arterial system is unnamed and largely undescribed. There have been some studies of the arteries based on casting and more recently CT imaging of the circulation of particular organs, such as the coronary circulation, and these will provide the data for our models. One of the reasons that there are so few named arteries is that the anatomy of the smaller arteries is highly variable. There are a number of publications in the surgical literature describing the distribution of variations in the named arteries and the variation is surprisingly large. [give some examples??]. Description of the unnamed arteries must rely on arterial models if their effect on arterial hemodynamics is to be studied.Another assumption used in almost all models of the arterial tree is that the tree is a uniform binary tree. This is a very convenient assumption because there is only one way to 'make' a uniform binary tree with \(N\) internal nodes. Analysis of the statistics of all possible trees with \(N\) internal nodes indicate that the 'average' tree is very different from a uniform binary tree. There are parallels here to the common perception that a three dimensional random walk will be approximately spherical in shape, which is by true for the average of many individual random walks. Detailed analysis of the statistics of a single random walk, however, give a very different picture. The three principle axes of the moment of inertia of a random walk have the ratios (11.7, 2.7, 1) which is closer to a slightly flattened cigar than a sphere. Unfortunately little is known about the 'average' binary tree, but we will try to incorporate what is known into our model.Probably the most famous study of blood vessel morphology is that of Murray (1926)\cite{1,Murray_1926,Murray_1926a} He analysed steady flow in a single bifurcation by minimising a cost function that included the hydraulic work needed to drive the flow assuming Poiseuille flow and the metabolic work needed to produce and sustain the blood within the vessels. He concluded that the diameters of the three vessels would have the relationship\(D_a^3=D_b^3+D_c^3\)where subscript a indicates the parent vessel and b and c indicate the two daughter vessels. This relationship is generally known as Murray's Law although, given the scatter observed in experimental test of the relationship, Murray's Hypothesis would probably be a better name. For an historical view of work in this area the background section of \cite{16143652} is highly recommended.The most recent extension of this work is the ZKM model.\cite{10616146} This model considers the vascular tree to be made up of stem-crown segments and applies a similar cost function minimising the combination of a steady Poiseuille flow hydraulic work and the metabolic work based on the blood volume in the entire tree. The minimisation analysis produce the relationships for the volume V, diameter D, flow rate Q and length L\(\frac{V}{V_{\max}}=\left(\frac{L}{L_{\max}}\right)^{\frac{5}{\epsilon+1}}\), \(\frac{D}{D_{\max}}=\left(\frac{L}{L_{\max}}\right)^{\frac{3\epsilon-2}{4\left(\epsilon+1\right)}}\), \(\frac{Q}{Q_{\max}}=\left(\frac{D}{D_{\max}}\right)^{\frac{4\left(\epsilon+1\right)}{3\epsilon-2}}\)where \(D_{\max},\ Q_{\max},\ V_{\max}\) and \(L_{\max}\) are the diameter and flow rate of the most proximal stem, the volume of the entire crown and the cumulative arterial length of the crown. The parameter \(\epsilon\) relates to the resistance of the crown and depends on the ratio of the metabolic to hydraulic work in the cost function. This model has been applied to a large number of experimental measurements from humans and other mammals and the results are generally good in that the model can be used to fit a very wide range of data.\cite{16143652} As the author points out, hoowever, the perception of the goodness of fit of the model is coloured by the range of the data plotted (the main figure is a log-log plot with 11 and 35 decades on the two axes) where quite large deviations of the fit would be barely visible. This work certainly advances our understanding of the structure of the arterial system.Both the Murray model and the ZKM model are based on the principle of minimum work and use the assumption that the flow in the arteries can be described by steady Poiseuille flow, a very strong assumption given the unsteady and complex nature of arterial flow. From a more fundamental point of view the basic assumption that the cardiovascular system is designed to optimise some relatively simple cost function is questionable. For example, it is impossible to explain the surprisingly high mean arterial pressure if the hydraulic work of the heart is taken into account. Most of the work done by the left ventricle comes from the necessity to raise left ventricular pressure to the level of the diastolic arterial presure so that the aortic valve will open. The fact that left ventricular pressures are approximately three times the right ventricular pressure is direct evidence that the high arterial pressure is not not necessary to blood flow necessary to perfuse the body since both sides of the heart produce the same cardiac output. One of the few logical explanations for high systemic arterial pressure is the need to perfuse the heart itself, which occurs predominantly during diastole when the myocardium is relaxing. This important feature of the cardiac system cannot be captured by a simple principle of minimum work.The evolutionary pressures that have shaped the design of our body can be very subtle. For example, adaptability is an important factor in evolution. In terms of the cardiac system this means that we have to be able to chase down our dinner even though we spend most of our time sleeping. Thus we have evolved the ability to more than double our cardiac output very quickly in response to specific cues. A cardiovascular sytem that was highly optimised for the inactive state would probably not be optimal in an evolutionalry sense even though it would undoubtedly result in a minimisation of our net hydraulic work. Our motivation in this work is to explore other models of the arteries that do not rely on the minimisation of a particular cost function.Theoretical backgroundAlthough the cardiological system may not necessarily obey the principle of minimum work, it does have to obey the basic laws of physics. In particular arterial hemodynamics must obey the basic laws of conservation of mass and momentum. We will make extensive use of these principes in the following work. Before we do that, however, we have to make some simplifying assumptions about the nature of arterial flow. The first assumption we will make is that the arteries can be described as a binary tree with the bifurcations being the nodes and the arterial segments between these nodes assumed to be uniform elastic tubes. Detailed analysis of arterial geometry indicates that arterial segments exhibit a small degree of taper, although many studies of arterial taper neglect the existence of small branches which result in an apparent decrease in the diameter of the main artery. For the sake of simplicity our model will assume that the arterial segments between bifurcations are uniform.We first derive the equations for flow in a single vessel. This is followed by a brief derivation of the conditions to be imposed at the nodes, bifurcations and terminal nodes. Finally we discuss the properties of rooted, signed trees and what we can learn about the arterial tree from the mathematical study of trees.1.1 flow in a single vesselWe assume the the edges in the tree can be modelled as uniform elastic tubes (arterial segments) whose properties vary with time and the axial distance along the artery (the 1-D approximation). Conservation of mass and momentum yield the partial differential equations for pressure P, local mean velocity U and local cross-sectional area A\(A_t+\ \left(UA\right)_x=0\)\(U_t\ +UU_x\ =-\frac{1}{\rho}P_x\ -\ \frac{8\pi\mu}{\rho}\frac{U}{A}\)where the subscipts \(t\) and \(x\) indicate partial differentials. The last term on the rhs of the second equation represents the viscous drag on the walls of the vessel due to viscosity assuming Poiseuille flow. To close the problem we assume that the elasticity of the vessel can be described by a tube law \(A=A\left(P\right)\) that assumes that the area of the tube is only a function of the pressure. This is an approximation that excludes viscoelatic behaviour, but has proven to be a reasonable assumption for the large arteries. The validity of this assumption in the small arteries is unknown.Using the tube law to eliminate \(A\) from the conservation equations we find that the equations are hyperbolic and can be solved using the method of characteristics.\cite{Parker_2009} This analysis provides a definition of the square of the local wave speed \(c^2\ =\ \frac{A}{\rho\frac{dA}{dP}}\) where \(\frac{dA}{dP}\) is the total derivative of the tube law with respect to pressure and is a measure of the compliance of the vessel. There are two comments about the wave speed that will be relevant to the ensuing analysis. In general the wave speed will be a function of presssure. The tube law that gives a constant wave speed can be found by the solving \(\frac{dA}{dP}=\frac{A}{\rho c^2}\) with \(\rho c^2\) constant and the boundary condition \(A\left(P_0\right)=A_0\): \(A=A_0e^{\frac{P-P_0}{\rho c^2}}\). For any other tube law \(c=c\left(P\right)\) although we commonly assume that \(c\) is constant in our analyses.The second comment is that it is possible to find an asymptotic expression for the area as \(c\ \rightarrow\ \infty\). For the general expression for the wave speed \(\frac{dA}{dP}=\frac{A}{\rho c^2}\)we can show by separating the variables and integrating that \(\ln\left(\frac{A}{A_0}\right)=\int_{P_0}^P\frac{dP'}{\rho c^2}\). Expanding this expression in an asymptotic series we see that \(A\ =\ A_0+O\left(\frac{1}{c^2}\right)\) in the limit of large \(c\). This is not surprising since rigid tubes have a constant area and an infinite wave speed.It is convenient to make use of the definition of wave speed in rewriting the conservation equations to the form\(P_t+UP_x+\rho c^2U_x=0\)\(U_t+\frac{1}{\rho}P_x+\ UU_x=\frac{8\pi\mu}{\rho}\frac{U}{A}\)The method of characteristics makes use of the eigenvalues of the matrix of coefficients of the x-derivative terms which are\(\lambda_{\pm}=U\ \pm c\)Along the characteristic directions \(\frac{dx}{dt}=\lambda_{\pm}=U\ \pm c\) the conservations equations can be written using the equation for the total derivative \(\frac{dY}{dt}=Y_t+Y_x\frac{dx}{dt}\) in the form of the two ordinary differential equations\(\frac{dU}{dt}\ \pm\frac{1}{\rho c}\frac{dP}{dt}=-\frac{8\pi\mu}{\rho}\frac{U}{A}\ \)These equations have been used extensively in the study of the the large arteries where the term on the rhs is usually neglected or incorporated into a numerical solution procedure.For the study of the behaviour of elastic vessels over the wide range of sizes and mechanicial properties found in the arterial tree it is convenient to non-dimensionalise the conservation equations. For each variable we can define the equivalent non-dimensional variable, denoted by a prime, by choosing an appropriate characteristic value for that parameter, denoded by a hat. For example, we define the non-dimensional velocity \(U'=\frac{U}{\hat U}\). where \(\hat U\)is chosen to be a velocity that is characteristic of the problem. Non-dimensionalisation is more of an art than a science and there are usually several ways to choose the characteristic values, some more convient than others. If we choose \(\hat{x}=\hat D\) to be a reference diameter of the tube and \(\hat U\) to be some characteristic mean velocity in the tube (usually dictated by the boundary conditions), then it is most convenient to chose \(\hat t=\frac{\hat D}{\hat U}\). Because both the area and the wave speed are functions of pressure it is convenient to define \(A_0=A\left(P_0\right)\) and \(c_0=c\left(P_0\right)\) as the area and wave speed at some reference pressure \(P_0\) and use these reference values to non-dimensionalise the area and pressure, \(A'=\frac{A}{A_0}\)and \(c'=\frac{c}{c_0}\). It is natural to take \(\hat{D}=D_0\) where \(A_0=\frac{\pi}{4}D_0^2\) is the diameter of the tube at the reference pressure. There are several choices for the characteristic pressure. For the analysis of pressure-flow waves in the tube the most convenient choice is \(\hat P=\rho c\hat U\). When waves are not important the most convenient characteristic pressure is realated to the dynamic pressure \(\hat P=\rho\hat U^2\). In the bulk of this work we will use the wave-based non-dimensional pressure \(P'=\frac{P}{\rho c_0\hat{U}}\) but will occasionally use the dynamic pressure-based non-dimensional pressure \(P''=\frac{P}{\rho\hat U^2}\). We note that \(P'=mP''\) where \(m=\frac{\hat{U}}{c_0}\) is the Mach number based on the reference wave speed in the vessel. With these choices for the characteristic values, the non-dimensional conservation equations are\(P'_{t'}+U'P'_{x'}+\frac{c'^2}{m}U'_{x'}=0\)\(U'_{t'}+\frac{1}{m}P'_{x'}+U'U'_{x'}=-\frac{32\ U'}{Re\ A'}\)where \(Re=\frac{\rho D_0\hat{U}}{\mu}\) is the Reynolds number based on the characteristic diameter. The non-dimensional equation for the characteristics is \(\frac{dx'}{dt'}=U\pm\frac{c'}{m}\). Along these two characteristic directions the non--dimensional conservation equations can be written in terms of the total time derivative\(\frac{dP'}{dt'}-\left(U'\pm\frac{1}{m}\right)P'_{x'}+U'P'_{x'}+\frac{1}{m}U'_{x'}=0\)\(\frac{dU'}{dt'}-\left(U'\pm\ \frac{1}{m}\right)U'_{x'}+\frac{1}{m}P'_{x'}+U'U'_{x'}=\frac{32\ U'}{Re\ A'}\)Simplifying the equations and adding and subtracting the first equation from the second wearrive at the method of characteristics solution of the original partial differential equations. On \(\frac{dx'}{dt'}=U'\pm\frac{1}{m}\)\(\frac{d}{dt'}\left(U'\pm P'\right)=-\frac{32\ U'}{Re\ A'}\)If there is no dissipation the rhs of this equation is zero and we the simple result that \(U'\pm P'\) are constant on the forward and backward characteristics respectively. Since the characteristic dirctions depend on the local velocithy the the determination of \(U'\left(x',t'\right)\) and \(P'\left(x',t'\right)\) is not always easy, but it is straightforward. If viscous effects are not negligible it is generally necessary to resort to numerical methods to solve particular problems. Most numerical codes used for solving the problems are based on this reduction of the partial differential equations to ordinary differential equations along the characteristic directions. In this study we will seek an analytical asymptotic solution that is valid in the limit of small Mach number \(m\ll1\).asymptotic solution for small Mach numberFor \(\ m\ll1\) we assume that the non-dimensional velocity can be written as \(U'=U'_0\left(1\ +\ mu\right)\) where \(U'_0\) is constant and \(u\) is of order 1. We also assume that the non-dimensional area can be written as \(A'=A'_0\left(1+ma\right)\) where \(A'_0\) is a constant. This assumption is justified by the above discussion about nearly rigid vessels. Substituting into the ODE along the forward characteristic direction \(\frac{dx'}{dt'}=U'+\frac{1}{m}\)\(\frac{dP'}{dt'}=-\frac{32}{Re}\ \frac{U'_0\left(1+mu\right)}{A'_0\left(1+ma\right)}-\frac{mdu}{dt'}\ \approx-\frac{32\ U'_0}{Re\ A'_0}+\mathcal O\left(m\right)\)This equation is valid along the forward characteristic and so we can write \(\frac{dP'}{dt'}=\left(U'+\frac{1}{m}\right)\frac{dP'}{dx'}=\frac{1}{m}\left(1+mU'_0\left(1+mu\right)\right)\frac{dP'}{dx'}\). Thus, to \(\mathcal O\left(m\right)\)\(\frac{dP'}{dx'}=-\frac{32mU'_0}{Re\ A'_0}\)We can also write the x-derivative along the forward characteristic \(\frac{dP'}{dx'}=P'_{x'}+\frac{1}{\left(U'+\frac{1}{m}\right)}P'_{t'}=P'_{x'}+\frac{m}{\left(1+mU'_0\left(1+mu\right)\right)}P'_{t'}\approx P'_{x'}+\mathcal O\left(m\right)\). Finally, recalling the alternative non-dimensionalisation of the pressure based on the dynamic pressure we can write, to \(\mathcal O\left(m\right)\)\(\frac{dP''}{dx'}=-\frac{32\ U'_0}{Re\ A'_0}\)where the derivative is now the derivative wrt the x'-axis. This is the non-dimensional equation for the relationship between pressure gradient and the mean velocity in Poiseuille flow; steady, viscous flow in an infinite rigid tube of circular cross-section.This analysis indicates that flow in the individual vessels of the arterial tree transitions from wave-dominated flow in the large arteries where \(Re\gg1\) to viscous-dominated flow when \(Re\ll1\) and \(m\ll1\). For the purpose of finding a function-based model of the arterial circulation this suggests that it may be necessary to consider different models for the large and small arteries.-----------------------------------For a large artery with diameter of order 1 cm and mean velocity of order 20 cm/s, the Reynolds number based on the mean velocity is of the order 1000 and so it is legitamate to neglect the term on the rhs. However for a small artery with diameter of order 1 mm and mean velocity of order 2 cm/s, the Reynods number is of order 0.05 and the viscous dissipation is no longer negligible. This simple dimensional analysis indicates that viscous dissipation is not only not negligivle but can become dominant in more distal vessels in the arterial tree.These two equations, one for each characteristic direction can be used to solve the 1-D problem and this is the general method of characteristic solution for \(P\) and \(U\) in a single elastic vessel. The solution for a network of uniform vessels can be found by applying the appropriate mathching conditions at the nodes. This method of solution will fail when the dissipation term on the rhs of the ODE becomes large. We can, however, extend the solution into this region by considering the solution of the ODE along the forward characteristic. Because the equation contains both dependent variables there are many possible solutions. One solution which is comppatible with the boundary conditions is\(U'\ =\ U'_0,\ \ \ \ \ \ \ \ \ \frac{dP'}{dt'}=\mp\frac{32\ U'_0}{Re\ A'_{ }}\)where \(U'_0\) is a constant given by the velocity at the origin of tube. Because the area is related to the pressure through the tube law, which can be rather complex, it is not possible to find a generally valid solution of this equation. However, if the tube is not very distensible we can set \(A'\ \approx A'_0\) to order \(m\). Along the forward characteristic we can write the derivative in terms of \(x\)\(\frac{dP'}{dx'}=\frac{-32}{ReA_0}\left(\frac{U_0}{U_0+\frac{1}{m}}\right)\ \approx\frac{-32\ m\ U_0}{Re\ A_0}\left(1-mU_0\right)\)In rigid tubes the wave speed is infinite and the approapriate scale for the pressure is the Bernoulli pressure. If we define the rescales non-dimensional pressure \(P''=\frac{P}{\rho U^2}=\frac{1}{m}P'\), the equation for the pressure gradient along the forward characteristic becomes, to order \(mU'_0\)\(\frac{dP''}{dx}=\frac{-32\ U_0}{Re\ A_0}\)which is the familiar form of the non-dimensional equation for flow through a rigid pipe. Furthermore, the x-derivative along the forward characteristic to the derivative long the x-axis to order \(m\); \(\frac{df}{dx}=\frac{\partial f}{\partial x}+\frac{1}{\frac{dx}{dt}}\frac{\partial f}{\partial t}=\frac{\partial f}{\partial x}+\frac{m}{1+mU}\frac{\partial f}{\partial t}\).1.2 conditions at nodesThe conditions in the vessels meeting at a node can be are describe by Kirchof conditions derived by applying the conservation of mass and momentum at the node. At a bifurcation we denote the parent vessel as \(a\) and the two daughter vessels as \(b\) and \(c\), and require\(Q_a=Q_b+Q_c\) and \(P_a=P_b=P_c\)In the wave-like arteries it is convenient to consider the dynamics of an incremental wavefront across which the pressure changes by \(dP\) and the velocity by \(dU\). Note that the changes can be either positive or negative. These waves are particularly simple and it is easy to build up any waveform as a series of sequential wavefronts. If the dissipative term is negligible, the method of characteristics solution gives rise to the water hammer equations relating \(dP\) and \(dU\)\(dP\ =\pm zdU\)where \(z\ =\ \frac{\rho c}{A}\) is the characteristic impedance of the vessel. The plus sign is for waves travelling tn the forward direction with speed \(U+c\) and the minus singn is fo waves travelling in the backward direaction with speed \(U-c\).If we introduce a wavefront in the parent vessel with magnitude \(\Delta P_a\), when it reaches the node it will generate a backward wavefront in the parent and two forward wavefronts in the two daughter vessels, denoted by \(dP_a,\ dP_b\) and \(dP_c\). Applying the Kirchof conditions and using the water hammer equations we find the reflection coefficient at the bifurcation\(\gamma_a=\frac{dP_a}{\Delta P_a}\ =\frac{y_a-y_b-y_c}{y_a+y_b+y_c}\)where we have defined the characteristic admittance \(y=\frac{1}{z}\). The transmission coefficient giving the magnitude of the wave transmitted into the other vessels is simply \(1+\gamma_a\). The reflection and transmission coefficients for waves approaching the node in the other two vessels can be found by cycling the subscripts. We note that the reflection and transmission coefficients are different for waves approaching the node in different different vessels, which will be an important later in the development of the model.Flow in the dissipative vessels is different in nature and is dominated by the resistance of the vessels in the network. For relatively rigid vessels we can write \(P_x=-RQ\) where \(R=\frac{4\kappa\mu}{A^2}\) is the resistance of the vessel. Note that the square of the area in the denominator say that the pressure drop depends on the fourth power of the diameter, which is typical of Poiseuille-like flows. Because trees are parallel in nature it is conveninet to define the admittance of the vessel \(S=\frac{1}{R}\). Then the admittance of a tree, or sub-tree, is simply the sum of all of the admittance downstream of the vessel \(\bar S=\Sigma S_i\) where the bar indicates the sum over all vessels downstream of the vessel.We can derive the reflection coefficient for a wave-like vessel joind to a tree of dissipative vessels by a similar calculation based on the Kirchov conditions at the not\(\Gamma_a=\frac{y_a-\bar S_b}{y_a+\bar S_b}\)where \(b\) refers to the dissipative vessel at the root of a tree of dissipative vessels. There is no transmission coefficient for this case since the dissipative vessels do not support waves. The reflection coefficients at a bifurcation where one daughter is a dissipative vessel and both daughters are dissapative vessels are\(\gamma_a=\frac{y_a-y_b-\overline{S}_c}{y_a+y_b+\bar S_c}\) and \(\gamma_a=\frac{y_a-\bar S_b-\bar S_c}{y_a+\bar S_b+\bar S_c}\)1.3 the properties of binary treesBinary trees are a particularly simple form of a network and it is convenient to use the nomenclature of graph theory to describe them. Each vessel corresponds to an edge and each edge has a node at either end. The number of edges connected to a node is defined as its degree. In binary trees nodes are either internal nodes of degree 3 or external nodes of degree 1. For the arterial tree external node connected to the ventricle is designated as the root. In a rooted binary the nodes can be unambiguously designated as the input and the output node depending on its proximity to the root. For a binary tree the total number of nodes is equal to the number of edges plus 1, \(N=E+1\). The number of external nodes is equal to the number of internal nodes, \(N_e=N_i+2\) and so \(E=2N_i+1\). Some authors refer to the internal nodes of a rooted tree simply as the nodes and the external nodes as the root and the leaves. Since we will be considering trees composed of two different types of edges, wave-like and dissipative, we will generally use internal and external nodes although it may sometimes be convenient to refer to the terminal nodes that connect to the microcirculation as the leaves. The height of a tree is defined as the number of edges between the root and the most distant terminal node.Because of their simplicity many models of the arterial tree assume that, at least at some level, the tree is a uniform binary tree. That is, at each generation each node is an internal node so that at each generation \(n\) there are \(2^g\) nodes with \(g=0\) being the root. Thus a uniform binary tree with \(g\) generations will have \(N_i=2^g-1,\ N_e=2^g+1\) and \(E=2^{g+1}-1\). The height of a uniform binary tree is equal to its numbr of generations, \(H=g\).Since most arterial bifurcations are asymmetrical it is most appropriate the think of it as a signed binary tree where the two daughter vessels are labelled major or minor. This is equivalent to decision trees where the two branches are lavelled, usually left and right. Because of the importance of decision trees in the design and search of data bases in computer technology, there has been a large amout of work on the properties of rooted, signed binary trees. An important property is the number of different trees there are. The number of distinct rooted, signed binary trees with \(n\) internal nodes is given by the Catalan number\(C_n=\frac{1}{n+1}\frac{\left(2n\right)!}{n!n!}\)which increases combinatorially as \(n\) increases. The Catalan numbers for \(n=0,1,2,3,4,5,6,7,8,9,10\) are \(1,1,2,5,14,42,132,429,1430,4862\) and \(C_{20}=1767263190\). The number of possible arterial trees is truely astronomical. A useful result for our work is that the average height of binary trees with \(n\) internal nodes is \(\bar H_n\sim2\sqrt{\pi n}\) as \(n\ \rightarrow\infty\).\cite{Flajolet_1982}The most convenient way to represent a tree mathematically is the connectivity matrix: an \(N\times E\) matrix where each row has a -1 in the column corresponding to its inlet node and a +1 at its outlet node. The degree of each node is simply the sume of the absolute values of the rows. The adjacency matrix, another common representation of a graph, is an \(\ E\times E\) matrix with a 1 in each column representing an edge that is connected by a node to the edge represented by the row. The adjacency matrix does not contain any information about the directivity of the tree and can be derived from the connectivity matrix. The labelling of the nodes and edges of a tree is arbitrary. One convenient convention for rooted trees which we will use is to label the root as 0, the edges in any convenient way with the outlet node of the edge being label the same as the edge. In trees with a loop this cannot be done because there will be one node that is the output node of two different edges. In this case the connectivity matrix can still be used but the choice of input and output nodes is arbitrary.1.4 self-similar treesA self-similar tree is a tree where each bifurcation obeys the same scaling law. The most general way to represent self-similar trees is using the formulation of L-systems (short for Lindenmayer systems, originally conceived as a mathematical theory of plant development).\cite{Prusinkiewicz_1990} L-system algorithms are powerful and apparently the method of choice for generating virtual reality plant life. However, they theory of L-systems is rather limited and so we will resort to simple self scaling laws.We assume that at each node there is a fixed ratio between the properties of the two daughter vessels and the parent vessel. In particular we assume that the diameter and lengths are given by\(\frac{D_b}{D_a}=\delta,\) \(\frac{D_c}{D_b}=\nu,\) \(\frac{D_c}{D_a}=\nu\delta\)\(\frac{L_b}{L_a}=\lambda,\) \(\frac{L_c}{L_b}=\mu,\) \(\frac{L_c}{L_a}=\mu\lambda\)where subscripts \(a,\ b,\ c\) refer to the parent and the major and minor daughters and all of the parameters are less than 1. The parameters \(\delta\) and \(\lambda\) represent the scaling between generations and \(\nu\) and \(\mu\) represent the asymmetry of the bifurcation. We will also assume that the other properties of a vessel are related to its diameter and length. The dependency of wave speed on diameter is not well-established but we will assume that \(c\ \sim D^{-\sigma}\).\cite{21622820} Other properties of interest are the characteristic impedance and the wave travel time\(z=\frac{\rho c}{A}\ \sim D^{-\left(2+\sigma\right)}\) and \(T=\frac{L}{c}\ \sim LD^{\sigma}\)Substituting the characteristic impedance into the equtions for the reflection coefficients at a node yield the scaling laws\(\gamma_a=\frac{1\ -\delta^{2+\sigma}-\left(\nu\delta\right)^{2+\sigma}}{1+\delta^{2+\sigma}+\left(\nu\delta\right)^{2+\sigma}}\), \(\gamma_b=\frac{\delta^{2+\sigma}-1-\left(\nu\delta\right)^{2+\sigma}}{1+\delta^{2+\sigma}+\left(\nu\delta\right)^{2+sigma}}\), \(\gamma_c=\ \frac{\left(\nu\delta\right)^{2+\sigma}-1-\delta^{2+\sigma}}{1+\delta^{2+\sigma}+\left(\nu\delta\right)^{2+\sigma}}\)In a self-similar tree these reflection coefficients, and their corresponding transmission coefficients, are the same at every internal node.