AUTHOREA
Log in
Sign Up
Browse Preprints
LOG IN
SIGN UP
Charles Pepe-Ranney
Postdoctoral fellow
Ithaca, NY
Member of:
Cornell University
Public Documents
3
The effect of carbon subsidies on marine planktonic niche partitioning and recruitmen...
Charles Pepe-Ranney and Ed Hall
Non-cyanobacterial diazotrophs mediate dinitrogen fixation in biological soil crusts...
Charles Pepe-Ranney, Chantal Koechli, Erin Eggleston, et al.
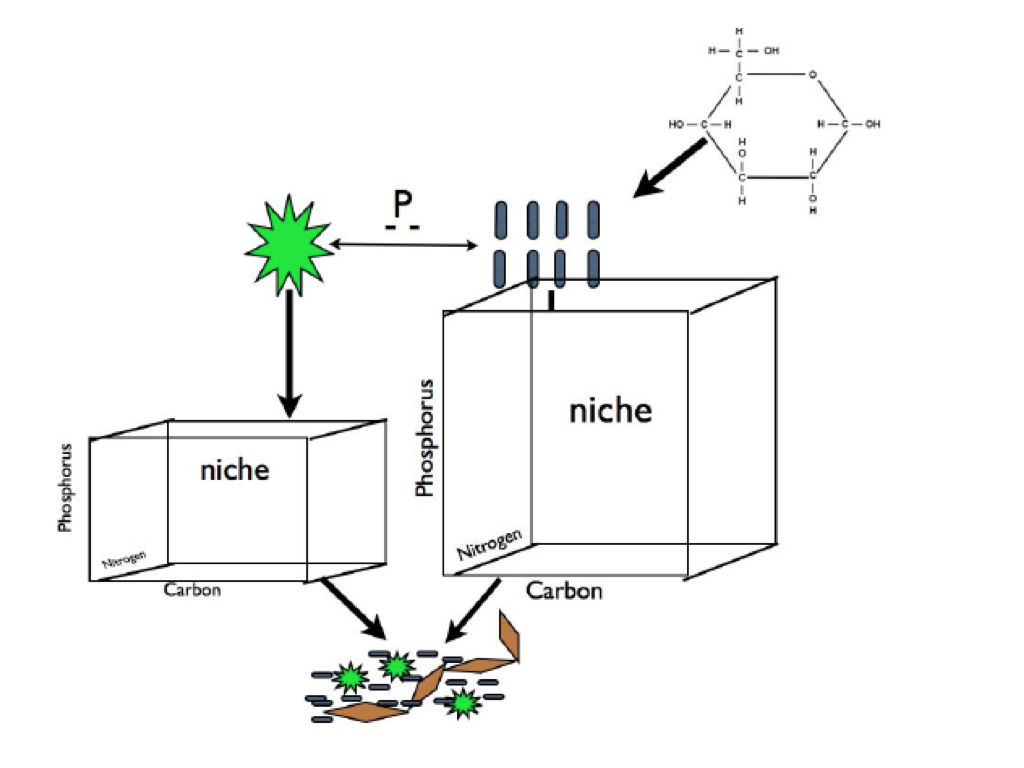
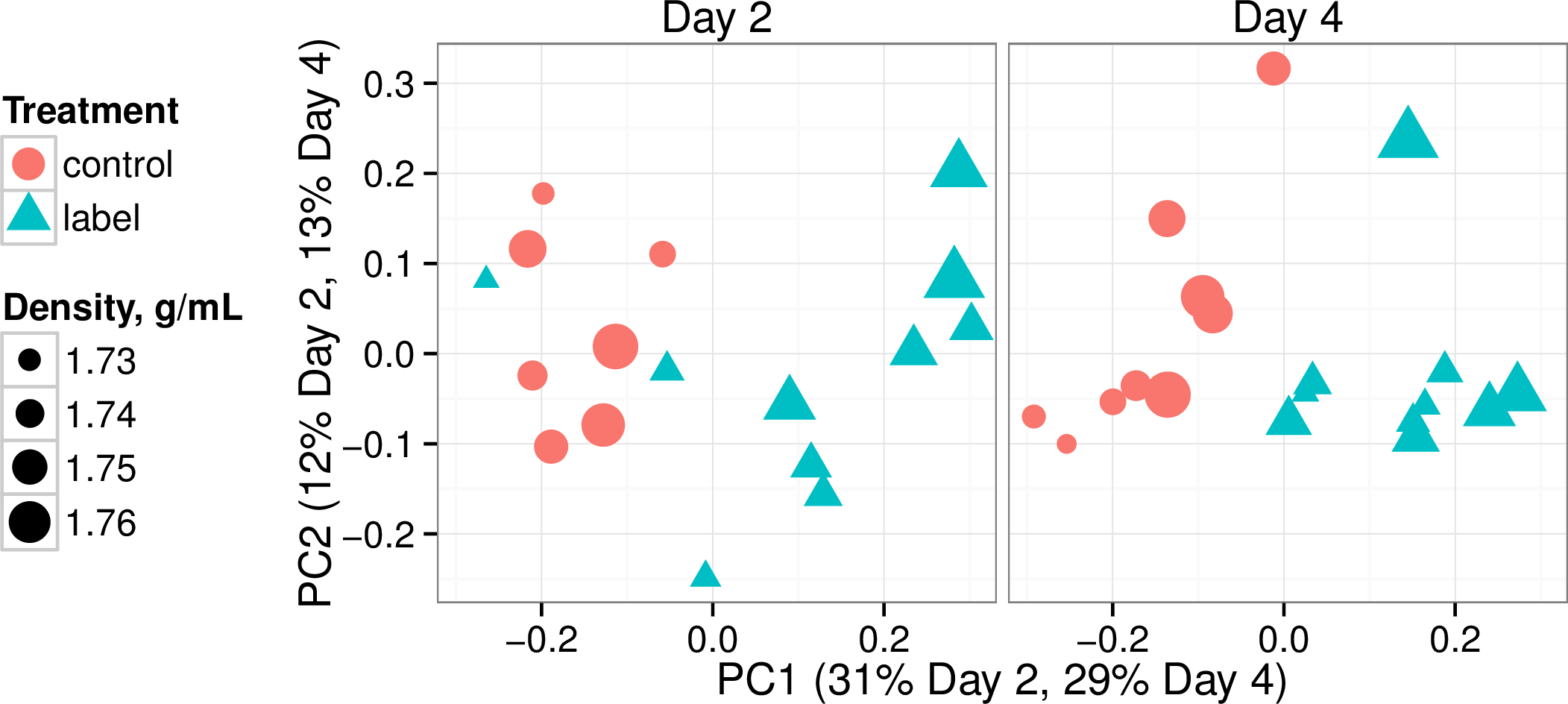
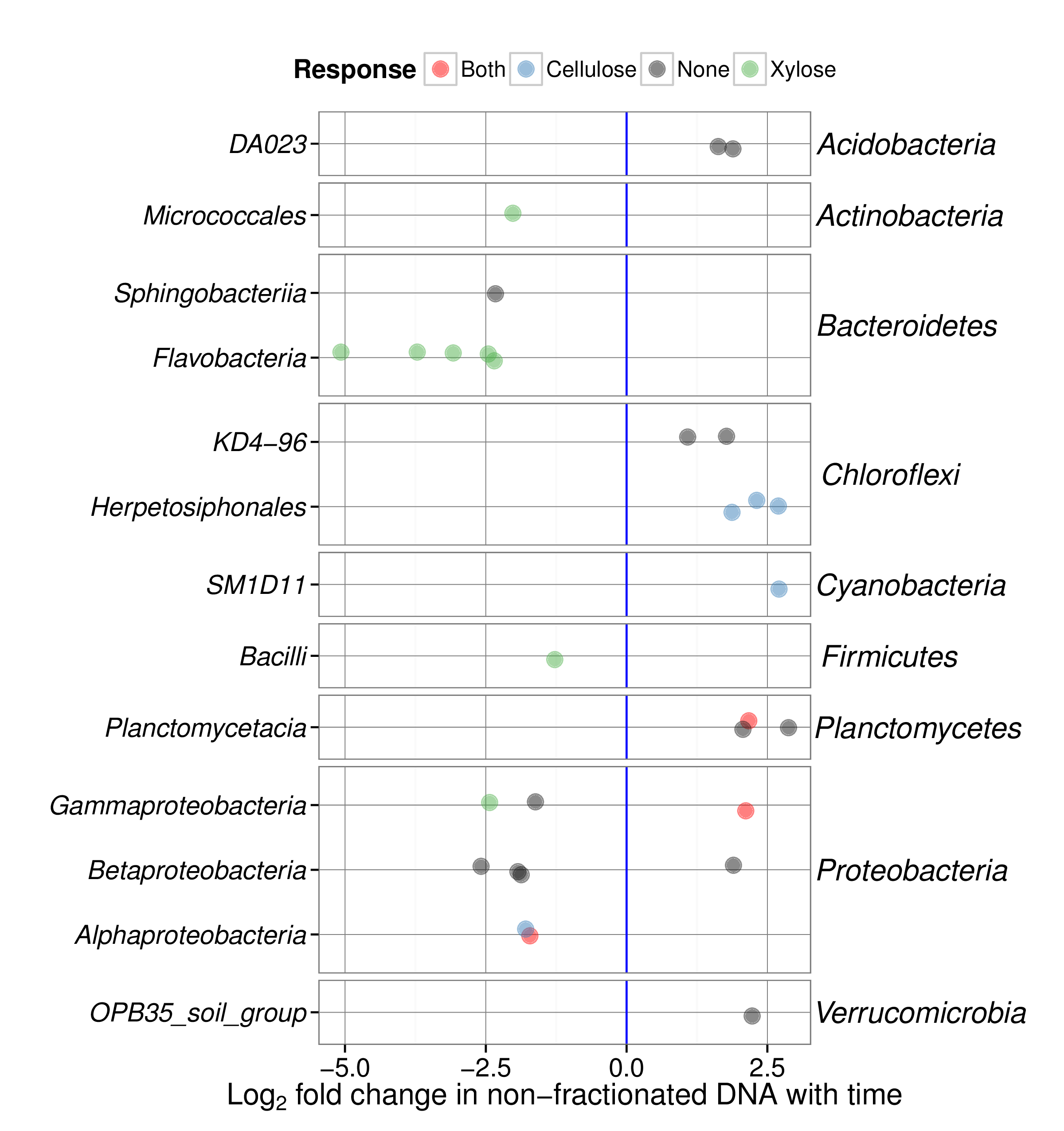
Unearthing the contributions and ecology of uncultured soil bacteria to the metabolis...
Ashley Campbell, Charles Pepe-Ranney, Dan Buckley, et al.