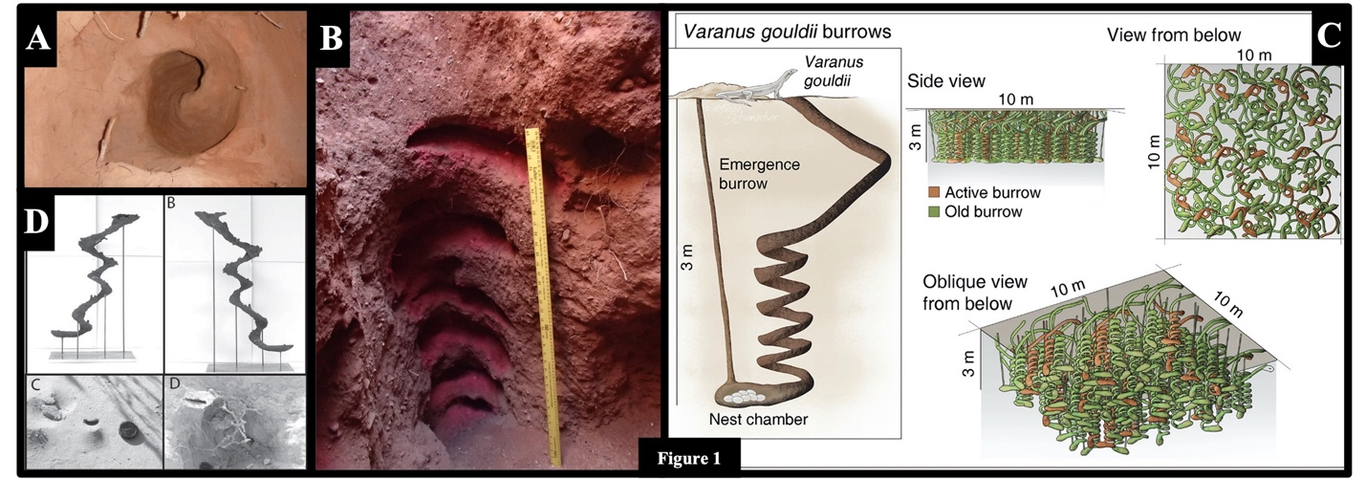
The extended phenotype of helical burrowing behavior in animals has evolved independently many times since first appearing after the Cambrian explosion (~540 million years ago). A number of hypotheses have been proposed to explain the evolution of helical burrowing in certain taxa, but no study has searched for a general explanation encompassing all taxa. We reviewed helical burrowing in both extant and extinct animals and from the trace fossil record and compiled from the literature 10 possible hypotheses for why animals construct helical burrows, including our own ideas. Of these, six were post-construction hypotheses—-benefits to the creator or offspring, realized after burrow construction—-and four were construction hypotheses reflecting direct benefits to the creator during construction. We examined the fit of these hypotheses to a total of 21 extant taxa and ichnotaxa representing 59–184 spp. Only two hypotheses—-antipredator, biomechanical advantage—-could not be rejected for any species (possible in 100% of spp.), but six of the hypotheses could not be rejected for most species (possible in 86–100 % of spp.): microclimate buffer, reduced falling sediment (soil), anticrowding, vertical patch, and the above two hypotheses. Four of these six were construction hypotheses, raising the possibility that helical burrowing might have evolved without providing post-construction benefits. Our analysis showed that increased drainage, deposit feeding, microbial farming, and offspring escape could not explain helical burrowing behavior in the majority of taxa (5–48%). Overall, the evidence does not support a general explanation for the evolution of helical burrowing in animals. The function and evolution of the helix as an extended phenotype would seem, at least in some cases, to provide different advantages for different taxa. Although direct tests of many of the hypotheses would be difficult, we nevertheless offer ways to test some of the hypotheses for selected taxa.